Autores
Alencar do Nascimento, D. (ESCOLA PROFESSORA CÂNDIDA ANDRADE MACIEL) ; Gomes da Silva, D. (UFPE) ; de Barros Corrêa, A.C. (UFPE) ; Albuquerque Meira Coelho Ramos, D. (UFPE) ; Calegari, M.R. (UNIOESTE)
Resumo
Nos estudos de reconstrução ambiental, o entendimento dos processos que causam as diferentes distribuições de corpos silicosos no solo é essencial para a interpretação dos fatores ambientais responsáveis por sua estabilidade, deposição de sedimentos, formação de paleossolos e análise de sítios arqueológicos. A extração e análise dos fitólitos foram obtidas nos Laboratórios de Análise de Formações Superficiais e Microscopia Ótica da UNIOESTE – Campus Francisco Beltrão. As conclusões a respeito da evolução ambiental para a área de Fazenda Nova foram baseadas na análise fitolítica dos depósitos das marmitas de dissolução. As análises da assembleia fitolítica indicaram que na área de Fazenda Nova o ambiente sempre possuiu uma vegetação aberta, apontando para condições ambientais quentes e secas causando stress hídrico, marcando a silicificação de células da epiderme de gramíneas.
Palavras chaves
Fitólito; Semiárido; Reconstrução Ambiental
Introdução
Nos estudos de reconstrução ambiental, o entendimento dos processos que causam as diferentes distribuições de corpos silicosos no solo é essencial para a interpretação dos fatores ambientais responsáveis por sua estabilidade, deposição de sedimentos, formação de paleossolos e análise de sítios arqueológicos. Os fitólitos, ou opala biogênica e/ou silicofitólitos, são corpos de sílica amorfa (SiO2.nH2O) produzidos pelas plantas ao longo dos seus ciclos vegetativos, e possuem dimensões semelhantes às da fração areia muito fina e silte do solo (CARTER 2002). As principais acumuladoras sílica e produtoras de fitólitos são as Poaceae, mas outras famílias de monocotiledôneas e algumas dicotiledôneas também acumulam quantidades expressivas de sílica (RUNGE 1999). Por serem as formas e ornamentações da sílica biogênica taxonômicamente únicas (CARTER op.cit.), possibilita que corpos silicosos de solos e sedimentos sejam usados na reconstituição de paleovegetação e paleoclimas. Além disso, os corpos silicosos podem ser usados na determinação da idade de camadas de sedimentos ou horizontes de solos, pois o carbono ocluso neles é preservado, principalmente em ambientes redutores (COSTA et al. 2010). Embora, algumas pesquisas tenham associado assembleia fitolítica à pólens na reconstrução da paleovegetação do Quaternário, a diferença entre os processos de formação, transporte e sedimentação destes taxas precisam ser considerados para uma melhor compreensão e corroboração dos dados apresentados. No Brasil, poucas pesquisas têm sido realizadas quanto às investigações referente à identificação de corpos silicosos em solos e sedimentos para fins de interpretação da gênese do solo e reconstrução dos cenários paleoambientais. Alexandre & Meunier (1999) estudaram a assembleia de fitólitos em latossolos do limite da floresta úmida e cerrado no município de Salitre (MG) e constataram uma fase de expansão da vegetação de savana entre 5.500 e 4.500 anos AP, associado a clima mais seco no Holoceno médio, enquanto dois períodos representados por comunidades arbóreo foram desenvolvidos entre 4.000 e 3.000 anos AP e a associação cerrado/floresta ocorrendo entre ~970 anos AP. Segundo os autores, essa tendência teria sido interrompido a partir de 970 ± 60 anos AP com a ocorrência de uma fase seca atual. Calegari (2008) constatou nos latossolos do cerrado de Salinas (MG) que a assembleia fitolítica era característica de mistura de plantas C3 e C4, sendo maior a contribuição de plantas C3, caracterizando uma vegetação aberta evidenciando mudança da vegetação e das condições de umidade ao longo do Holoceno na área. A ocorrência elevada de fitólitos de palmeiras e de poaceae com cores amareladas seriam indícios da ocorrência de paleoincêndios em torno de 6.000 e 4.000 anos AP, corroborando os estudos de Alexandre & Meunier (op.cit.) quanto à ocorrência de climas mais secos na região Sudeste no Holoceno médio. Entretanto, ao tratar do ecossistema semiárido, Costa et al. (2010) constataram a pobreza de sílica biogênica dos latossolos e vertissolos nordestino sob remanescente de floresta subperenifólia, caatinga hiperxerófila arbustiva e campos antrópicos em Granhuns (PE), Cachoeirinha (PE) e Crato (CE). Ramírez et al. (2007) chamam a atenção para o fato de que altos níveis de evapotranspiração em ambiente quente e úmido favorece a precipitação de ácido monossilícico na epiderme foliar da planta no lugar do fitólito. Do contrário, os ambientes mais secos a absorção do ácido monossilícico seria irrelevante, pois somente uma pequena concentração de sílica estaria disponível para a planta, explicando assim, a quase ausência de fitólitos em ambientes semiáridos. Sendo assim, a análise de assembleia de fitólitos é uma ferramenta acurada para caracterizar comunidades de vegetação dominada por poaceae que nem sempre são detectados pela análise polínica e isotópica, e para diferenciação de vegetação aberta (campos/padrarias) da vegetação mais fechada de florestas
Material e métodos
A extração e análise dos fitólitos foram obtidas nos Laboratórios de Análise de Formações Superficiais e Microscopia Ótica da UNIOESTE – Campus Francisco Beltrão, conforme Madela et al. (1998). Para a extração de fitólitos foram usados 4 gramas de solo/sedimento, que foram submetidos a uma “limpeza” para eliminação dos carbonatos com HCl, da matéria orgânica com peróxido de hidrogênio (33%) e eliminação da fração argila com Calgon. A concentração dos fitólitos foi realizada com Politungstato de Sódio (Sometu®) com densidade 2,35 e coletado com pipeta pasteur. A fração coletada foi seca em estufa 75oC e pesada para estimar a quantidade da fração ácida insolúvel na qual se encontram os fitólitos. A classificação dos morfotipos seguiu o ICPN 1.0 (MADELLA et al. 2005), os significados taxonômico e ambiental foram definidos a partir de Twiss et al. (1969), Mulholland (1989), Fredlund e Tieszen (1994), Alexandre et al. (1997; 1999), Runge (1999), Parr e Watson (2007) e Raitz (2012). Devido à limitação da análise fitolítica em termos de redundância (mesmo morfotipo produzido por variadas plantas) e multiplicidade (produção de vários morfotipos por uma mesma planta) de morfotipos (ROVNER 1971), agrupou-se os morfotipos mais representativos dentro dos grandes grupos taxonômicos já conhecidos para complementar à interpretação da vegetação como: bilobate e cross (Panicoideae), saddle (Chloridoideae), rondel (Pooideae)e bulliform, elongate e hair (Poaceae), block (carenate, irregula, polyedric e facetet), irregularplane, globulares rugose e psilate (eudicotiledonea), globularechinate (Arecaceae) e hexagonalcyperus (Cyperaceae). Para identificar as características da vegetação, tais como a dominância de subfamília de gramíneas (Poaceae), a densidade de cobertura arbórea e as condições de umidade, aplicou-se, respectivamente, os seguintes índices fitolíticos: a) cobertura arbórea (D/Po); b) climático (Ic); c) aridez (Iph); e d) índice de stress hídrico( FS%). Os índices foram calculados usando-se os morfotipos de fitólitos com significado taxonômico relevante: - D/Po estabelece a relação entre fitólitos diagnósticos de plantas arbóreas (eudicotiledonea) tropicais (globular granulate) e fitólitos de células curtas diagnósticas de gramíneas (Poaceae = rondel + trapeziform + polylobate + trapeziformshort cell + saddle + cross + bilobate short cell) (BREMOND et al. 2005). - Ic corresponde à porcentagem de fitólitos de células curtas, produzidas principalmente por Pooideae (rondel + trapeziform polylobate + trapeziform short cell), em relação a todas as células curtas produzidas pela soma de gramíneas Pooideae, Chloridoideae e Panicoideae (trapeziform polylobate + trapeziform short cell + saddle + cross + bilobate short cell) (TWISS 1992). - Iph estabelece a razão entre a porcentagem de fitólitos de células curtas de Chloridoideae (sanddle) e a soma dos morfotipos de gramíneas Panicoideae e Chloridoideae (sanddle + cross + bilobate short cell) multiplicado por 100 (DIESTER-HAAS et al. 1973, TWISS 1992). -Bi índice buliforme é a razão entre os morfotipos bulliformes (paralelepipedal bulliform e cuneiform bulliforme) e a soma de todos os fitólitos de gramíneas com significado taxonômico,exceto os elongates (sanddle + cross + bilobate short cell+trapeziform short cell+ trapeziforme polylobate) (DEHLON 2005) -FS índice de stress hídrico é a porcentagem do morfotipo fanshaped (Cuneiforme Bulliforme) em relação à soma de todos os fitólitos de gramíneas com significado taxonômico, exceto os elongates (sanddle + cross + bilobate short cell+trapeziform short cell+ trapeziforme polylobate) (BREMOND et al. 2005).
Resultado e discussão
Na área de Fazenda Nova, município de Brejo da Madre de Deus – PE (Figura 01),
a sedimentação quaternária encontra-se, em sua maioria, associada à ocorrência
de marmitas de dissolução e rampas coluviais sendo estes os principais
compartimentos de estocagem de sedimentos nas paisagens semiáridas aqui
consideradas.
A primeira área de coleta situa-se em um amplo anfiteatro de terras baixas
circunscritos a sul por um lineamento de serras graníticas que se elevam a mais
de 1.000m no município de Brejo da Madre de Deus, Pernambuco. O interior da
depressão apresenta um relevo aplainado e parcialmente dissecado circunscrito
pela isolinha de 500 m. Neste setor, além dos pedimentos e rampas de baixa
declividade, ocorrem também inselbergs, ambos dominados por uma cobertura
superficial de neossolos litólicos e planossolos. Em torno dos inselbergs
encontram-se comumente uma série de depressões fechadas produzidas pelo ataque
da erosão diferencial e remoção dos mantos de intemperismo, as marmitas de
dissolução.
A análise de uma assembleia fitolítica permite caracterizar uma formação
vegetal. Calibrações entre assembléias fitolíticas, fisionomia das vegetações e
limites climáticos mostram que os fitólitos são marcadores confiáveis e
precisos das vegetações e principalmente das diferentes formações herbáceas.
Sendo assim, foram analisadas amostras das marmitas de dissolução das
Propriedades Pocinho D'Água em Fazenda Nova e João Ferreira em Afrânio, a fim
de se identificar as características da vegetação, tais como a dominância de
subfamília de gramíneas (Poaceae), a densidade de cobertura arbórea e as
condições de umidade. Também foram analisados sedimentos da marmita de
dissolução da propriedade Incó, igualmente localizado no distrito de Fazenda
Nova, datadas por Silva & Corrêa (2009), para uma melhor caracterização
paleoambiental da área.
Na seção estratigráfica da propriedade Pocinho D'Água observou-se baixa
frequência e variedade de fitólitos quando comparados com solos e sedimentos de
outras regiões e biomas do Brasil (ALEXANDER et al. 1997; BORBA–ROSCHEL et al.
2006; CALEGARI et al. 2011). As amostras MP30/1A e MP130/1A não apresentaram
fitólitos, sendo estas consideradas estéreis para silicofitólitos, assim como
para outros corpos silicosos, como espículas de esponjas diatomáceas, muito
presentes nas demais amostras analisadas (Tabela 01 e Figura 02).
A assembleia fitolítica das amostras indica presença de fitólitos de Poaceae
(gramíneas), Arecaceae e Eudicotiledoneas (arbustos/arvores). Na amostra
MP70/1A, 73% da assembleia é formada por fitólitos de Poaceae, sendo somente 2%
de fitólitos com significado taxonômico, isto é, 1% de Bilobate e Cross
(Panicoideae) e 1% Pooideae (Rondel), sendo o restante, Bulliform (28%),
Elongate (35%) e Hair (8%) que são produzidos por praticamente todas as
gramíneas, e partes das gramíneas, não apresentando, desta forma, significado
taxonômico, apenas ambiental. Os fitólitos de arbustos e árvores (Eudicot -
globularspsilate e rugose e polyedricblock) representam 9% da assembleia dessa
amostra e os fitólitos de Arecaceae (globularechinate) representa 15% do
conjunto identificado.
Na amostra MP90/1A observou-se a mesma tendência, com os fitólitos de Poaceae
dominando o conjunto identificado (77%), sendo as gramíneas de padrão
fotossintético C4 as dominantes entre os fitólitos com significado taxonômico,
isto é, 3% de Panicoideae (Bilobate e Cross), 2% de Chloridoideae e apenas 1%
de Pooidea (Poaceae C3). Os fitólitos de arbustos e arvores representam 22% da
assembleia e os de Arecaceae 12%.
O morfotipo globular echinate pequeno (<6 µm) e escuro foi encontrado em todas
amostras, representando as palmeiras da família Arecaceae presente na área.
Cecchet (2012) avaliando a produção fitolítica de várias espécies dessa família
presentes na Floresta Ombrófila Densa de Linhares (Espírito Santo), encontrou
fitólitos idênticos em várias espécies, com hábitos ecológicos variados (Figura
03).
De modo geral, os índices fitolíticos indicam que o ambiente sempre possuiu uma
vegetação muito esparsa, aberta. Os fitólitos classificados como de
Eudicotiledoneas, são, predominantemente, os blocos poliédricos (polyedric
block) identificados em espécies arbustivas em coleções de referencias
elaboradas no Brasil para os Biomas Campo e Floresta Ombrófila Mista com
Araucária no Paraná (RAITZ, 2012) e em menor frequência em espécies arbóreas da
Floresta Ombrófila – Mata de Tabuleiro no Espírito Santo (LIMA, 2012).
Sendo assim, os índices apontam para condições ambientais quente e seca (IC
3,2% e 2,3%) a moderadamente úmida (Iph 1,4%), com momentos de seca causando
stress hídrico conforme indicado pelos valores do Índice de stress hídrico que
marcam a silicificação de células da epiderme de gramíneas como resposta a
escassez de água em algum período do desenvolvimento da planta ao longo do
tempo. Essa tendência é reforçada pelo Índice de stress hídrico (FS%) cujos
valores variaram entre 11,5 e 3,5 nas amostras MP70/1A e MP90/1A,
respectivamente. De acordo com Bremond et al. (2005), quanto mais a planta
transpira e/ou sofre stress hídrico, mais células buliformes silicificadas ela
poderá produzir.
De acordo com os baixos valores do índice D/P (<0,1) não houve uma vegetação
fechada, ou que chegasse a formar um dossel fechado, haja vista que estes
valores são muito baixos e indicam uma vegetação aberta, formada por espécies
de gramíneas e arbustos.
Juntamente com os fitólitos, foi encontrado grande quantidade de fragmentos de
diatomáceas e de espículas de esponja que indica ter havido uma lamina de água,
ou água em quantidade suficiente para permitir o desenvolvimento desses
palinomorfos.
Em todas as amostras desta seção os sinais de corrosão foram observados na
maioria das partículas (morfotipos de fitólitos, fragmentos de diatomáceas e de
espículas, grãos de minerais), sobretudo naquelas de maior tamanho como nas
bulliforms e blocos (menor superfícies específica). Isso indica se tratar de um
ambiente cujos valores de pH podem ser extremos, ou muito baixo (<2,0) ou muito
altos (>7,5), pois os ambientes alcalinos são os que mais atacam a sílica, seja
biogênica ou não. Essas condições explicariam também a baixa
quantidade/frequência de fitólitos células curtas (saddle, bilobates, rondel e
cross), pois como possuem elevada superfície específica, são os primeiros a se
degradarem ao longo do tempo.
Para as amostras da propriedade Incó observou-se que a diversidade de
morfotipos é baixa (Tabela 02 e figura 04). Foram identificados apenas seis
morfotipos, sendo eles: os bilobates (Panicoideae), bulliform, hair e
elongates(Poaceae). Estes representam mais de 60% da assembleia de cada amostra
estudada, sendo os hexagonais (Cyperaceae) e irregular block (Eudicotiledonea)
entre 20 e 25% das assembleias.
Essa baixa diversidade indica predomínio de fitólitos de monocotiledôneas C4, e
uma vegetação aberta, conforme corroborada pelos valores nulos do índice de
cobertura arbórea. Coleções de referencia elaboradas com espécies de outros
biomas tem reconhecidos os morfotipos irregular blocks, polyedrics blocks em
várias espécies de arbustos. Como ainda não existem coleções de referencias com
espécies do bioma da Caatinga, é necessário cautela ao analisar a ocorrência
dos blocks nessas amostras.
Os dados apontam baixa cobertura arbórea (D/P 0), ainda que os fitólitos de
elementos arbustivos e arbóreos representem em torno de 25% da assembleia
identificada nas amostras Incó. Não foram encontrados fitólitos globulares
rugoses e psilates que são usados para calculo desse índice.
O stress hídrico e a alta evapotranspiração das plantas são bem marcados na
amostra superficial pelo índice Fs % (10%), principalmente quando comparado com
a amostra subjacente. O índice de aridez (Iph%) apontam para condições mésicas
no ambiente, o que parece não coincidir com as condições ambientais
representadas pela amostra de 0-10 cm.
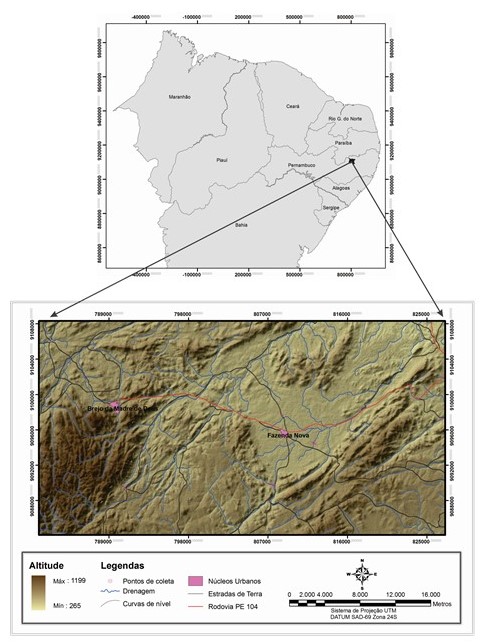
Localização da área de Estudo
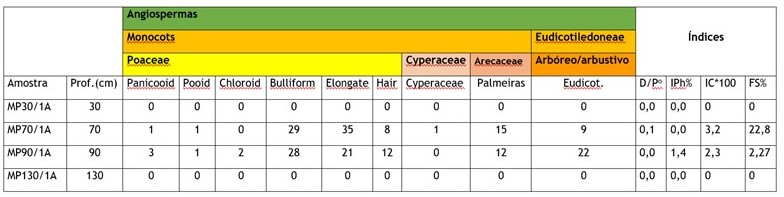
Porcentagem de cada morfotipo em relação à assembleia identificada e índices fitolíticos para o testemunho Pocinho D'Água.
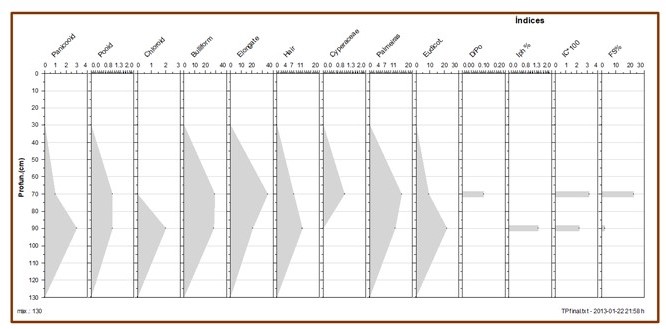
Diagrama de fitólitos (em porcentagem) para o testemunho Pocinho D'Água.
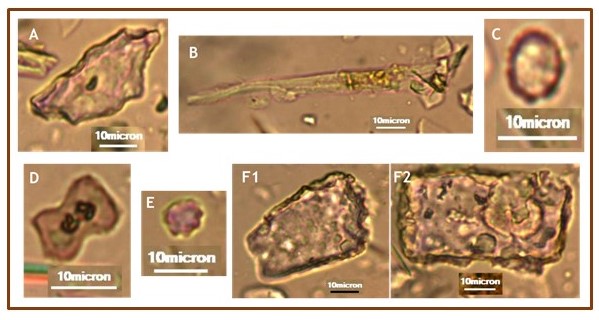
Principais morfotipos identificados:A) Irregular faceted block; B)acicular(hair);C) Globular psilate;D) Bilobate; Cross; E) Globular echinate;

Porcentagem da soma total de fitólitos identificados (com significado Taxonomico /ambiental) para o testemunho Incó.
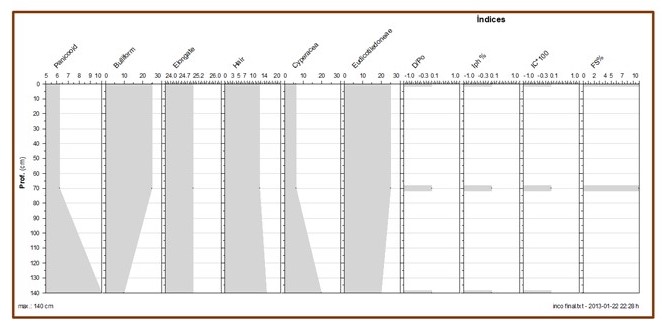
Diagrama de fitólitos (em porcentagem) para o testemunho Incó.
Considerações Finais
As conclusões a respeito da evolução ambiental para a área de Fazenda Nova foram baseadas na análise fitolítica dos depósitos das marmitas de dissolução. Os depósitos que preenchem as marmitas de dissolução foram derivados por sedimentação gravitacional do tipo fluxo de detrito, através de fenômenos de episódios espasmódicos durante chuvas torrenciais em clima semiárido. As análises da assembleia fitolítica indicaram que na área de Fazenda Nova o ambiente sempre possuiu uma vegetação aberta, apontando para condições ambientais quentes e secas causando stress hídrico, marcando a silicificação de células da epiderme de gramíneas. A interpretação da dinâmica geomorfológica através do registro sedimentar e fitolítica encontrado nas marmitas de dissolução forneceram bases para a elucidação de interações entre as mudanças temporais de longo e curto prazo nos processos geomorfológicos no Quaternário tardio na região, cujas repercussões ainda são visíveis na paisagem. Entretanto, a principal dificuldade dessa proposta incide na natureza intrínseca do material, pois os depósitos na forma que se encontram na paisagem, representam apenas uma pequena parcela dos materiais originais que conseguiram permanecer incólumes às perturbações decorrentes das mudanças ambientais da ordem de centenas a milhares de anos.
Agradecimentos
Referências
ALEXANDER, A.; MEUNIER, J.-D.;LÉZINE, A.-M.; VINCENS, A.; SCHWARTZ, D. Phytoliths indicators of grasslands dynamics during the late Holocene in intertropical Africa. Palaeogegraphy, Palaeoclimatology, Palaeocology v.136, p.213-219, 1997.
ALEXANDRE, A. & MEUNIER, J. -D. Late Holocene Phytolith and Carbon-Isotope Record from Latosol at Salitre, South-Central Brazil. Quaternary Research, 51, p. 187-194, 1999.
BRÉMOND, L., ALEXANDRE, A. ; HÉLY, C.; GUIOT, J. A phytolith index as a proxy of tree cover density in tropical areas: calibration with Leaf Area Index along a forest-savanna transect in southeastern Cameroon. Global and Planetary Changes, 45. 2005, pp. 277-293.
CALEGARI, M. R. Ocorrência e significado paleoambiental do horizonte A húmico em Latossolos. Tese de Doutorado. Escola Superior de Agricultura Luiz Queiroz, 2008. 259p.
CARTER, J. A. Phytolith analysis and paleoenvironmental reconstruction from
Lake Poukawa Core, Hawkes Bay, New Zealand. Global and Planetary Change, 33, p. 257–267, 2002.
CECCHET, F. Conjunto de fitólitos dos estratos inferiores da floresta ombrófila densa (Linhares – Espírito Santo): subsídios para reconstrução paleoambiental. Monografia de conclusão de curso – Curso de Geografia (Licenciatura) - Centro de Ciências Humanas, Educação e Letras, Universidade Estadual do Oeste do Paraná - UNIOESTE- Campus Marechal Cândido Rondon. 2012. 87 pg.
COSTA, L. M. et al. Ocorrência de corpos silicosos em horizontes superficiais de solos de diferentes ecossistemas. Rev. Bras. Ciências do Solo. 34, p. 871-879, 2010.
DELHON, C. Phytolith and pedoanthracology analysis of “off-site”Holocene sequences from Mandragon (Middle Rhone Valley, south of France). In: Zurro, D. (Ed.) Madella, M. Recent Studies in Phytolith Analysis, Oxbow books, Plants Peoples and Places, 2007. Pp.175-188.
DIESTER-HAASS, L., SCHRADER, H.J., THIEDE,J. Sedimentological and paleoclimatological investigations of two pelagic ooze cores off Cape Barbas, North- West Africa. Meteor Forshungergebnisse v.16, p.19-66, 1973.
FREDLUND, G. TIESZEN, L. T. Modern phytolith assemblage from the North American Great Plains. Journal of Biogeography v.21, p.321 -335, 1994.
LIMA, P. L. F. Coleção de referência de fitólitos de Eudicotiledoneae da Floresta Ombrófila Densa: subsídios para estudos paleoambientais em regiões tropicais. Monografia de conclusão de curso – Curso de Geografia (Licenciatura) - Centro de Ciências Humanas, Educação e Letras, Universidade Estadual do Oeste do Paraná - UNIOESTE- Campus Marechal Cândido Rondon. 2012. 62 pg.
Madella, M., Power-Jones, A., Jones, M., A simple method of extraction of opal phytoliths from sediments using a non-toxic heavyliquid. Journal of Archaeological Science 25, 801–803, 1998.
MADELLA, M.; ALEXANDRE, A.; BALL, T. International Code for Phytolith Nomenclature 1.0. Annals of Botany, Oxford, v. 96, p. 253–260, 2005.
MULHOLLAND, S.C. Phytolith shape frequencies in North Dakota grasses: a comparison to general patterns. Journal of Archeological Science, v.16, p.489-511, 1989.
RAITZ, E. Coleção de referência de silicofitólitos da flora do Sudoeste do Paraná: subsídios para estudos paleoambientais, 204 f. Dissertação (Mestrado em Geografia), Universidade Estadual do Oeste do Paraná, Campus Francisco Beltrão. 2012.
RAMIREZ, S. H. U. et al. Disolución de sílice biogênica em sedimentos de lagos utilizados como bioindicadores de calidad del água. Gnosis, 5, p. 1-19, 2007.
ROVNER, I. Potential of opal phytoliths for use in paleoecological reconstruction. Quaternary Research v.1, p.343-359, 1971.
RUNGE, F. The opal phytolith inventory of soils in Central Africa - Quantities, shapes, classification, and spectra. Rev. Paleobot., 107, p. 23-53, 1999.
SILVA, D. G. & CORRÊA, A. C. B. Evolução paleoambiental dos depósitos de tanques em Fazenda Nova, Pernambuco - Nordeste do Brasil. Rev. Bras. Geografia Física, v. 2(2), p. 43-56, 2009.